
Abstract
Discoloring biofilms from Cambodian temples Angkor Wat, Preah Khan, and the Bayon and West Prasat in Angkor Thom contained a microbial community dominated by coccoid cyanobacteria. Molecular analysis identified Chroococcidiopsis as major colonizer, but low similarity values (<95%) suggested a similar genus or species not present in the databases. In only two of the six sites sampled were filamentous cyanobacteria, Microcoleus, Leptolyngbya, and Scytonema, found; the first two detected by sequencing of 16S rRNA gene library clones from samples of a moist green biofilm on internal walls in Preah Khan, where Lyngbya (possibly synonymous with Microcoleus) was seen by direct microscopy as major colonizer. Scytonema was detected also by microscopy on an internal wall in the Bayon. This suggests that filamentous cyanobacteria are more prevalent in internal (high moisture) areas. Heterotrophic bacteria were found in all samples. DNA sequencing of bands from DGGE gels identified Proteobacteria (Stenotrophomonas maltophilia and Methylobacterium radiotolerans) and Firmicutes (Bacillus sp., Bacillus niacini, Bacillus sporothermodurans, Lysinibacillus fusiformis, Paenibacillus sp., Paenibacillus panacisoli, and Paenibacillus zanthoxyli). Some of these bacteria produce organic acids, potentially degrading stone. Actinobacteria, mainly streptomycetes, were present in most samples; algae and fungi were rare. A dark-pigmented filamentous fungus was detected in internal and external Preah Khan samples, while the alga Trentepohlia was found only in samples taken from external, pink-stained stone at Preah Khan. Results show that these microbial biofilms are mature communities whose major constituents are resistant to dehydration and high levels of irradiation and can be involved in deterioration of sandstone. Such analyses are important prerequisites to the application of control strategies.
Similar content being viewed by others


Introduction
The complex of temples associated with Angkor Wat, in Cambodia, were built mainly from gray sandstone by the Khmer people between the 8th and 13th centuries, AD. They include Angkor Wat itself, Angkor Thom, Preah Khan, and the Bayon, a massive temple that is best known for the bas-relief carving and Buddha faces on the towers. Further out at this site and set in an opening in the forest is West Prasat, which began as a Hindu shrine, perhaps built in the 10th century, but was reconstructed as a Buddhist temple from the 13th century.
The buildings are situated in a tropical climate with alternating intense dry and rainy seasons. The general regression rate of the temple buildings is 2.1–8.3 mm ka−1 [3], but biodeterioration causes increased losses. Higher plants are seen growing in many temples, lichens are rampant and cyanobacteria-dominated black biofilms, leading to thermal stress on the surfaces, are widespread [4]. There have been few investigations into microbial fouling at these sites. A German project initiated in 1995 showed that the microbial biofilms were complex yet stable communities of mainly superficial algae, fungi, and bacteria. An update can be found in Warscheid and Leisen, 2011 [38].
There has been no detailed study published of the bacteria and phototrophs in the biofilms, apart from the lichens [5]. Microbial phototrophs have been considered to be the most important primary colonizers of external structures [11] and cyanobacteria are major biofilm components and deteriogens of historic stone buildings in tropical climates [8]. Removing them from the stone surfaces of Angkor Wat temples left the structures open to attack by potentially more damaging fungi [37]. We aimed to make a study of the bacteria present on the buildings at the Angkor Wat site, in addition to noting the other microorganisms constituting these biofilms.
Materials and Methods
Sampling
Samples were taken from surface biofilms on buildings in Angkor Wat (AW), Preah Khan (PK) and, in Angkor Thom, the Bayon (B) and West Prasat (WP), using the non-invasive adhesive tape method [15]. About 10 cm samples were taken from various sites on each building, covering different biofilm macromorphologies. Figures 1, 2, and 3 show some of the sampling sites. Biofilms were green, gray, brown, black, and pink in color. Interior surfaces (Samples 1 from PK and B) were dark green or dark brown, and moister than external biofilms, which were most commonly gray or black. Lichen growth (on external surfaces) was not sampled to avoid detecting non-free-living phototrophs and fungi.

Gray and black biofilms on Angkor Wat. Some pink/red staining (possibly Trentepohliales) is seen on the left (arrow)

External gateway at Preah Khan, showing extensive mixed biofilms (green, pink/brown and gray), including white and pale green lichens. A pink/brown-stained area is arrowed. Pillars and ceiling within the structure (seen in the center of the photograph) carry thick, moist, deep green biofilms

Photograph taken from inside building at the Bayon, Angkor Thom, showing difference between moist green/brown internal biofilms, clearly seen above the window, and dry external gray films containing many white and pale green lichens
Microbiological Analysis
Samples were divided into approximately 2 cm strips, placed on solid algal/cyanobacterial media, MN [18], BG11 and MKM [15], and incubated under constant light conditions (80 μmol photons m−2 s−1) at 25°C. As detailed in Shirakawa et al. [15], they were examined microscopically after 1–4 h of rehydration, to determine major biomass in situ, and then inspected at intervals (daily for the first week) up to 8 weeks. At intervals, strips were moved to new plates to allow better growth of organisms which had transferred to the agar. Cyanobacteria were identified by morphology [6]; algae, fungi, and other bacteria, including actinomycetes, were recorded and morphology noted. Some isolated or mixed colonies picked from solid media and suspended in sterile saline, as well as three tape strip samples (approx. 5 cm washed into 1 ml) were used for molecular analyses.
Molecular Analyses
Total DNA was extracted from each sample [10]. It was used as template to amplify the V3 hypervariable region of bacterial 16S rRNA using the primers P3 (5′-CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGGCCTACGGGAGGCAGCAG-3′) and P2 (5′-ATTACCGCGGCTGCTGG-3′) [27]. The amplification mixture contained 100 ng DNA, 2.5 μl 10× reaction buffer, 50 mM MgCl2, 0.25 mM of each dNTP, 10 pmol of each primer, and 1 U Taq Polymerase (Invitrogen Corporation, Carlsbad, CA), adjusted to 25 μl. The PCR program was: 94°C for 5 min; 35 cycles of 1 min at 94°C, 1 min at 55°C, and 1 min at 72°C; and a final extension at 72°C for 5 min. The PCR products were applied to 8% (w/v) denaturing polyacrylamide gels and DNA bands visualized by silver staining [32]. Prominent bands were selected, excised and used as template for the PCR performed as described above. Bands were sequenced with P2 primer and a distance matrix was produced based on the Dice correlation coefficient.
Ribosomal 16S rRNA gene libraries were constructed using cyanobacterial primers designed from aligned sequences of at least one species of each cyanobacterial order and of other bacteria. One forward primer CYAFor (5′-AGCAGTGGGGAATTTTCC G-3′) and two reverse primers CYACRev (5′-GGTCAGCATACGGCRGTGAAT-3′), and CYAARev (5′-GGTCAGCATACTGCRGTGAAT-3′) were synthesized and used in the PCR at 1 pmol each with 20 ng template DNA. PCR products were purified, cloned into pJET1.2/Blunt using the GeneJET PCR Cloning Kit (Fermentas International Inc., Ontario, Canada) and the resulting ligation products used to transform chemocompetent Escherichia coli DH10b cells. Transformants were PCR screened directly for the presence of inserts using pJET1.2 vector primers (10 pM/reaction). Nucleotide sequences were determined using the pJET1.2 primers (Macrogen Inc, DNA Sequencing Service, Seoul, Korea) and sequences analysed by BLASTn through the Ribosomal Database Project-II Release 9 (http://rdp.cme.msu.edu). Alignments via CLUSTAL X were manually edited using SEAVIEW software [13]. Paired similarity and pairwise distance calculations using the transversion/transition weighting (R = s/v) and the Kimura-2-parameter model [21] were performed with the MEGA version 4.0 program. Phylogenetic trees were constructed using the neighbor-joining method with 1000 bootstrap replications. Similarity percentages were calculated using BioEdit 7.0.5.2 software. The genus and species limits were 95 and 97.5%, respectively [31]. The 26 new sequences were deposited in the GenBank with accession numbers 7CYA_2 (FJ985811), 7CYA_3 (FJ985812), 7CYA_4 (FJ985813), 7CYA_5 (FJ985814), 7CYA_6 (FJ985815), 8CYA_2 (FJ985816), 8CYA_7 (FJ985817), 9CYA_A8 (FJ985818), 9CYA_24 (FJ985819), 9CYA_25 (FJ985820), 9CYA_26 (FJ985821), 9CYA_27 (FJ985822), 9CYA_28 (FJ985823), 14CYA_27 (FJ985824), 14CYA_F5 (FJ985825), 14CYA_H6 (FJ985826), 14CYA_8 (FJ985827), CYA_Band_A (FJ985828), CYA_Band_B (FJ985829), CYA_Band_C (FJ985830), CYA_Band_D (FJ985831), CYA_Band_E (FJ985832), CYA_Band_F (FJ985833), CYA_Band_G (FJ985834), CYA_Band_H (FJ985835), and CYA_Band_I (FJ985836).
Results
Microscopy
The microorganisms detected microscopically in the biofilm communities are shown in Table 1. Although small, single-celled bacteria were seen in large numbers in all cases, cyanobacteria constituted the main biomass, as shown by direct examination of rehydrated biofilms. Subsections I and II organisms were predominant. Filamentous cyanobacteria were detected in only two samples, both from internal biofilms (B1 and PK1). Actinobacteria, mainly streptomycetes, were seen in most samples. Algae and fungi were rare. A dark-pigmented filamentous fungus was detected only in PK samples, and the green alga Trentepohlia only in samples taken from external, pink-stained areas of PK.
Molecular Analysis
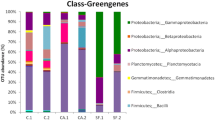
A total of seventeen 1100 bp nucleotide sequences from ribosomal libraries were analyzed. Members of the phylum Cyanobacteria were detected and are shown in Table 2. Figure 4 shows the neighbor-joining phylogenetic tree. Major bands from PCR–DGGE of samples and clones (Fig. 5; Table 2) showed the cyanobacteria Chroococcidiopsis sp. (subsection II) in AW and PK1 samples nos. 8, 9, and 14, Microcoleus sp. (subsection III) in PK1 sample no. 14 and various bacteria in AW, B1, PK1, and PK2 samples nos. 5, 7, 8, 9, and 11.

Neighbor-joining phylogenetic tree of 16S rRNA gene sequences cloned directly from metagenomic DNA of biofilms. Sequences were aligned with ClustalX, and distances were calculated with the Kimura 2-parameter substitution model. The scale bars indicate the nucleotide substitutions per site. Numbers at the branches indicate the bootstrap values from 1,000 resamplings. Only values above 50% are shown. Paenibacillus polymyxa and P. massieliensis served as outgroup

PCR–DGGE banding profiles of 16S rRNA bacterial genes from biofilms. Lanes 1–6 correspond to biofilm samples 5 (B1), 7 (PK2), 8 (AW), 9 (PK1), 11 (PK2), and 14 (PK1 tape sample); lanes 7–11 correspond to ribosomal library clones 14CYA_27, 14CYA_F5, 7CYA_2, 8CYA_2, 9CYA_24. Letters on left of lanes indicate bands that were excised for sequence analysis (Table 1). Arrows indicate profiles corresponding to library clones
Discussion
The detection of a principal biomass of cyanobacteria in the biofilms on buildings in the Angkor Wat complex mirrors the situation reported on stone buildings in India and Latin America [1, 14, 28]. Cyanobacteria are resistant to desiccation and high solar irradiation (15 and references therein). This obviously acts in their favor on stone surfaces in tropical countries. The predominance of coccoid subsection I and II organisms also agrees with results obtained in other countries [15, 28]. The molecular biology results indicate a prevalence of Chroococcidiopsis, a genus of subsection II, although the similarity values for species are mainly rather low (<95%). The DGGE data suggest that a single Chroococcidiopsis sp. was present, associated with heterotrophic bacteria. However, resistance of some cyanobacteria to lysis, lack of reaction with the primers used, and inhibition of the PCR by cell constituents, such as pigments, may have restricted detection of cyanobacterial genera [16, 29]. A comparison of partial 16S rDNA sequences of 22 cyanobacteria isolated from walls of Brazilian buildings with 52 sequences from data bases found that the isolates fitted well within their families, but that the similarity values were less than 96% [17]. The authors suggested that terrestrial cyanobacteria are a distinct population from the aquatic organisms whose DNA sequences constitute the majority of database deposits, emphasizing the need for polyphasic analyses of terrestrial biofilms. Low similarity values were also found for cyanobacterial clones in a molecular biology study of biofilms on the Bayon, Angkor Thom [24]; however, a wide range of cyanobacteria were detected, numerically the most common after Proteobacteria. Since no microscopic analyses were carried out, these results cannot be compared with the totality of ours.
Subsection II organisms were identified as major biomass in all biofilms by microscopy (Table 1) and the molecular results confirm this. The inability of the latter to identify the cyanobacteria more closely once more indicates the differences between these terrestrial cells and those currently in databases, as previously reported [16, 17, 36]. The subsection II organisms have previously been stated to be members of more established communities on historic buildings [9, 23, 30] and these Angkor Wat biofilms fit this description.
Filamentous cyanobacteria were only detected in moister biofilms. In PK1, a moist green biofilm from an internal wall in Preah Khan (see Fig. 2), filamentous cyanobacteria of subsection III, probably the Microcoleus detected by band sequencing from DGGE gels and identified as Lyngbya by microscopy (for problems with Microcoleus taxonomy see, for example, Ref. [7]), were among the major colonizers, with a possible Leptolyngbya detected via the gene library. Scytonema (Subsection IV) was detected in sample B1, taken from an internal pillar in the Bayon. These internal biofilms were much moister than external ones, being less subject to direct sunlight and air movements. An inspection of the literature suggests that filamentous cyanobacteria are much more common than coccoid in aquatic environments [2, 33, 35] and this difference in humidity probably explains the incidence on internal versus external walls.
A second difference between internal and external biofilm communities was the presence of the green alga Trentepohlia, found at PK. These cells were detected only in samples taken from the exterior, pink-stained areas of PK, confirming the typical stone discoloration caused by this organism and already reported in Edzna, Mexico [14], and Singapore [39]. In these pink areas, Trentepohlia was the major biomass. Pink-stained areas seen on other buildings suggested that this algal genus was not restricted to PK. This was, in fact, checked at some sites by wiping a humid finger across the wall and observing the change in color from pink to deep orange/red, typical of Trentepohliales biofilms.
Small single-celled bacteria were seen microscopically in all samples and were often spore-forming rods. Such microorganisms, the low GC Firmicutes, prominent in undisturbed and agricultural soils ([26] and references therein), have been detected in the biofilms on historic monuments by both traditional and molecular techniques [19, 20, 22, 25, 29]. DNA sequencing of excised DGGE bands indicated that members of this phylum were present in our biofilms, as well as two members of the Proteobacteria (Table 2). Several species of Bacillus and Paenibacillus were positively identified, together with Lysinibacillis fusiformis.
One sample, denominated number 11, which showed 100% similarity to the proteobacterium Methylobacterium radiotolerans, was a pink colony isolated from a plate incubated with an external PK sample. This is the first time that this facultatively methylotrophic bacterium has been detected on sandstone buildings. Like some Bacillus spp. and cyanobacteria, M. radiotolerans is resistant to high levels of irradiation, giving it an advantage over less tolerant organisms on exposed surfaces in tropical countries. Similar resistance is shown by dark-pigmented fungi, detected in PK. These fungi were also detected on the Bayon, Angkor Thom, using only DNA sequencing methods [24]; the same article confirmed our finding of low fungal presence overall.
The microorganisms detected in biofilm communities on the surfaces of stone monuments in the Angkor Wat temple complex, Cambodia, may contribute to the deterioration of these structures. Cyanobacteria, the major components of the community, are detrimental not only because of the surface discoloration produced by their pigments, but also through erosive metabolites and, in some cases, direct stone boring activity (8 and references therein). Microbial phototrophs are probably the first colonizers of stone and constitute a significant proportion of biofilm biomass; their mere presence can provide nutrients for heterotrophic microorganisms, which act as stone deteriogens through organic acid production or mechanical action. Certain paenibacilli (P. glycanilyticus and P. sepulcri) effectively degrade cyanobacterial polysaccharides, producing acids from these and other carbon sources [12, 34]. The surface communities on stone also lead to the production of nutrients and niches suitable for the development of higher plants, with their destructive root systems.
Knowledge of the microbial composition of fouling terrestrial biofilms is necessary for the development and monitoring of strategies to prevent biodeterioration and biodegradation of historic stone monuments.
References
Adhikary SP, Satapathy DP (1996) Toypothrix byssoidea (Cyanophyceae/Cyanobacteria) from temple rock surfaces of coastal Orissa, India. Nova Hedwigia 62:419–423
Anand N, Mohan E, Hopper RSS, Subramanian TD (1986) Taxonomic studies on blue green algae from certain marine environments. Seaweed Res Util 9:49–56
André MF (2006) Sandstone weathering rates at the Angkor temples (Cambodia). In: Fort R, Alvarez de Buergo M, Gomez-Heras M, Vasquez-Calvo C (eds) Heritage, weathering and conservation. Taylor & Francis, London
André MF, Etienne S, Mercier D, Vautier F, Voldoire O (2008) Assessment of sandstone deterioration at Ta Keo temple (Angkor): first results and future prospects. Environ Geol 56:677–688
Anon (2006) International coordinating committee for the safeguarding and development of the historic Site of Angkor. Report of Fifteenth Technical Committee—June 5, 6, 7, 2006, p 90
Boone DR, Castenholz RW, Garrity GM (2001) Bergey’s manual of systematic bacteriology, vol 1. Springer, New York
Boyer SL, Johansen JR, Flechtner VR, Howard GL (2003) Phylogeny and genetic variance in terrestrial Microcoleus (Cyanophyceae) species based on sequence analysis of the 16S rRNA gene and associated 16S–23S ITS region. J Phycol 38:1222–1235
Crispim CA, Gaylarde CC (2005) Cyanobacteria and biodeterioration of cultural heritage: a review. Microbial Ecol 49:1–9
Crispim CA, Gaylarde CC, Gaylarde PM (2004) Biofilms on church walls in Porto Alegre, RS, Brazil, with special attention to cyanobacteria. Int Biodeter Biodegr 54:121–124
Cullen DW, Hirsch PR (1998) Simple and rapid method for direct extraction of microbial DNA from soil to PCR. Soil Biol Biochem 30:983–993
Darlington A (1981) Ecology of walls. Heinemann, London
Dasman KajiyamaS, Kawasaki H, Yagi M, Seki T, Fukusaki E, Kobayashi A (2002) Paenibacillus glycanilyticus sp. nov., a novel species that degrades heteropolysaccharide produced by the cyanobacterium Nostoc commune. Int J Syst Evol Microbiol 52:1669–1674
Galtier N, Gouy M, Gautier C (1996) SEA VIEW and PHYLO_WIN: two graphic tools for sequence alignment and molecular phylogeny. Comput Appl Biosci 12:543–548
Gaylarde P, Englert G, Ortega-Morales O, Gaylarde C (2006) Lichen-like colonies of pure Trentepohlia on limestone monuments. Int Biodeter Biodegr 58:119–123
Gaylarde CC, Gaylarde PM (2005) A comparative study of the major microbial biomass of biofilms on exteriors of buildings in Europe and Latin America. Int Biodeter Biodegr 55:131–139
Gaylarde CC, Gaylarde PM, Copp J, Neilan BA (2004) Polyphasic detection of cyanobacteria in terrestrial biofilms. Biofouling 20:71–79
Gaylarde PM, Crispim CA, Neilan BA, Gaylarde CC (2005) Cyanobacteria from Brazilian building walls are distant relatives of aquatic genera. OMICS 9:30–42
Gaylarde CC, Ortega-Morales BO, Bartolo-Perez P (2007) Biogenic black crusts on buildings in unpolluted environments. Curr Microbiol 54:162–166
Heyrman J, Swings J (2001) 16S rDNA Sequence analysis of bacterial isolates from biodeteriorated mural paintings in the Servilia tomb (Necropolis of Carmona, Seville, Spain). Syst Appl Microbiol 24:417–422
Kiel G, Gaylarde C (2006) Bacterial diversity in biofilms on external surfaces of historic buildings in Porto Alegre. World J Microbiol Biotechnol 22:293–297
Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16:111–120
Laiz L, Piñar G, Lubitz W, Saiz-Jimenez C (2003) Monitoring the colonization of monuments by bacteria: cultivation versus molecular methods. Environ Microbiol 5:72–74
Lamenti G, Tiano P, Tomaselli L (2000) Biodeterioration of ornamental marble statues in the Boboli Gardens (Florence, Italy). J Appl Phycol 12:427–433
Lan W, Li H, Wang W-D, Katayama Y, Gu J-D (2010) Microbial community analysis of fresh and old microbial biofilms on Bayon Temple Sandstone of Angkor Thom, Cambodia. Microb Ecol 60:105–115
McNamara CJ, Perry TD, Bearce KA, Hernandez-Duque G, Mitchell R (2006) Epilithic and endolithic bacterial communities in limestone from a Maya archaeological site. Microbial Ecol 51:51–64
McSpadden Gardener BB (2004) Ecology of Bacillus and Paenibacillus spp. in agricultural systems. Phytopathology 94:1252–1258
Muyzer G, De Waal EC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695–700
Ortega-Morales O, Guezennec J, Hernandez-Duque G, Gaylarde CC, Gaylarde PM (2000) Phototrophic biofilms on ancient Mayan buildings in Yucatan, Mexico. Curr Microbiol 40:81–85
Ortega-Morales BO, Narvaez-Zapata JA, Schmalenberger A, Dousa-Lopez A, Tebbe CC (2004) Biofilms fouling ancient limestone Mayan monuments in Uxmal, Mexico: a cultivation-independent analysis. Biofilms 1:79–90
Peraza Zurita Y, Cultrone G, Sánchez Castillo P, Sebastián E, Bolívar FC (2005) Microalgae associated with deteriorated stonework of the fountain of Bibatauín in Granada, Spain. Int Biodeterior Biodegr 55:55–61
Rosselló-Mora R, Amann R (2001) The species concept for prokaryotes. FEMS Microbiol Rev 25:39–67
Sanguinetti CJ, Neto ED, Simpson AJG (1994) Rapid silver staining and recovery of PCR products separated on polyacrylamide gels. Biotechniques 17:914–921
Scheffer M, Rinaldi S, Gragnani A, Mur LR, van Nes EH (1997) On the dominance of filamentous cyanobacteria in shallow, turbid lakes. Ecology 78:272–282
Smerda J, Sedlácek I, Pacova Z, Krejcí E, Havel L (2006) Paenibacillus sepulcri sp. nov., isolated from biodeteriorated mural paintings in the Servilia tomb. Int J Syst Evol Microbiol 56:2341–2344
Valério E, Faria N, Paulino S, Pereira P (2008) Seasonal variation of phytoplankton and cyanobacteria composition and associated microcystins in six Portuguese freshwater reservoirs. Ann Limnol 44:189–196
Walker JJ, Pace NR (2007) Phylogenetic composition of rocky mountain endolithic microbial ecosystems. Appl Environ Microbiol 73:3497–3504
Warscheid T (2000) Integrated concepts for the protection of cultural artefacts against biodeterioration. In: Of microbes and art: the role of microbial communities in the degradation and protection of cultural heritage. Kluwer, Amsterdam, pp 185–201
Warscheid T, Leisen H (2011) Microbiological studies on stone deterioration and development of conservation measures at Angkor Wat. In: Charola AE, McNamara C, Koestler RJ (eds) Biocolonization of stone: control, preventive methods. Smithsonian Inst, Washington, pp 1–18
Wee YC, Lee KB (1980) Proliferation of algae on surfaces of buildings in Singapore. Int Biodeter Bull 16:113–117
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gaylarde, C.C., Rodríguez, C.H., Navarro-Noya, Y.E. et al. Microbial Biofilms on the Sandstone Monuments of the Angkor Wat Complex, Cambodia. Curr Microbiol 64, 85–92 (2012). https://doi.org/10.1007/s00284-011-0034-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00284-011-0034-y